El rabijunco etéreo (Phaethon aethereus) es un ave marina tropical que habita en áreas del Atlántico, del este del Pacífico y del noroeste del Índico. Está en declive a escala global. Se estima una población mundial de 3.000-13.000 ejemplares maduros y no califica como amenazada por la UICN (LC, Preocupación menor).
En Canarias es un colonizador reciente que se observa en época de cría por todo el archipiélago, no estando incluido en ninguno de los catálogos de especies protegidas, ni nacionales ni autonómicos. En los últimos años ha criado de forma muy escasa en El Hierro (3 parejas con cierta regularidad) y Lanzarote (1 pareja). En 2016 se localizó una colonia en Fuerteventura (20-25 individuos, 7-8 nidos) y es probable que haya una segunda colonia.
[ADDENDA. A final de abril de 2018, dentro de la 1ª Campaña del Proyecto Phaeton, se han marcado en Fuerteventura varios ejemplares con dispositivos GPS, adultos capturados en las cuevas donde tienen los nidos. Alguna fuente habla de 4 parejas presentes.]
Fenología en Canarias: inmensa mayoría de observaciones entre febrero y julio. Actividad reproductiva entre marzo (ocupan territorio) y julio (pollos abandonan el nido). Hay cierto desfase respecto a Cabo Verde (unas 1.000 parejas), donde está presentes todo el año y crían desde diciembre. Por cercanía, Cabo Verde es el origen más probable de los ejemplares de Canarias, aunque no puede descartarse su origen en el Caribe.
lunes, 30 de abril de 2018
domingo, 29 de abril de 2018
ESCRIBANO PALUSTRE EN ESPAÑA. CENSO AÑO 2015
El escribano palustre cuenta
con dos subespecies reproductoras en la península ibérica: el escribano
palustre iberoriental (E. s. witherbyi), que se reproduce en el interior, en
la costa mediterránea española y francesa y en la costa atlántica de Marruecos;
y el escribano palustre iberoccidental (E. s. lusitanica), reproductor en la
cornisa cantábrica y atlántica de la península ibérica. Ambas son sedentarias y
figuran como “En Peligro” en el Libro Rojo de las Aves de España publicado en
2004.
En 2005 se realizó el I Censo Nacional en España. Con resultado: E. s. witherbyi 250-360 parejas, la mayoría en Castilla-La Mancha; E. s. lusitanica 62-68 parejas, mayoría en Galicia.

Diez años después, en 2015, tras constatar un empeoramiento, se realizó con voluntarios el II Censo Nacional y I Censo Ibérico, coordinado por la Universidad de Valencia y la Universidad Politecnica de Valencia. El censo fue completo en España, pero no en Portugal, se visitaron 180 humedales (24 en Portugal, de los 31 potenciales), incluyendo todas las localidades censadas en 2005. En España, la población total es de 155-170 parejas (319-431 en 2005), con un declive del 50-60% en 10 años, por lo que la especie debería ser considerada en España y Portugal como En Peligro Crítico B2b (i,ii,iii,iv,v) según los criterios de la UICN.
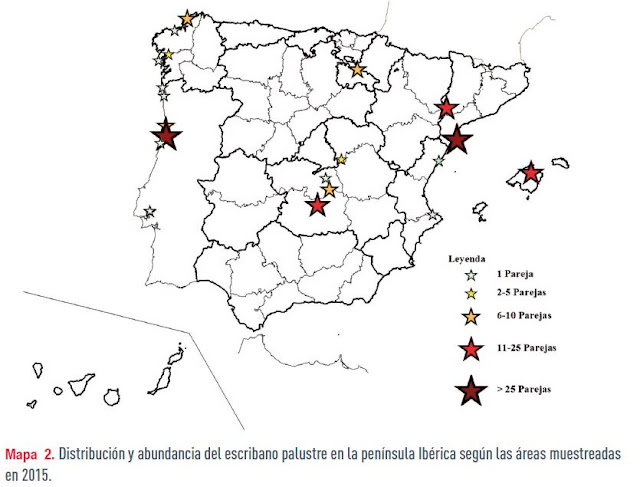
El censo de escribano palustre iberoccidental fue de 96-101 parejas en 11 humedales, 5 en España (13 en 2015) y 6 en Portugal. En Portugal sólo se muestrearon el 77% de las localidades potenciales. La mayor parte se concentra en una localidad, Salreu, ría de Aveiro, Portugal (75-77 parejas). En España sólo hubo ejemplares en Galicia, con un censo de 11-12 parejas y un declive del 83% respecto a 2005 (un censo posterior, en 2018, localizó 20 parejas en Galicia). Para Portugal no existe un censo previo para comparar, aunque se detectó un descenso en el número de humedales con presencia como reproductor.
Para el escribano palustre iberoriental, el censo fue de 144-158 parejas reproductoras en 10 localidades (21 en 2005). El humedal con mayor población fue el Delta del Ebro (63-65%), seguido de Castilla-La Mancha (17-16%) y el P. N. de S’Albufera de Mallorca, única localidad de las islas Baleares (15-13%). Respecto a 2005 se ha detectado un declive del 43-56%, siendo más severo en las Tablas de Daimiel donde en 2005 se estimaron 90-120 parejas frente a las 15 detectadas en 2015. El escribano palustre iberoriental se reproduce en 2015 en sólo 10 humedales (21 en 2005) de Castilla-La Mancha, Cataluña, Comunidad Valenciana, Baleares y Navarra. En el litoral mediterráneo es estable en el Delta del Ebro, se reduce en la Comunidad Valenciana y aumenta en S’Albufera de Mallorca.

Monrós, J. S., Neto, J. M., Arcos, F., Vera, P. y Belda, E. J. 2018. El escribano palustre en España y
Portugal. Población reproductora en 2015 y método de censo. Universitat de València-Universitat
Politécnica de València-SEO/BirdLife. Madrid. https://doi.org/10.31170/0059
LOCALIDADES POSITIVAS EN ESPAÑA, 2005 y 2015
Laguna de los Ojos de Villaverde (Albacete) 7 - 0
Laguna de Manjavacas (Cuenca) 2 - 0
Embalse de Buendía-Río Mayor (Cuenca) 3 - 0
Complejo Lagunar de Arcas (Cuenca) 2 - 0
Embalse de Almoguera (Guadalajara) 7 - 3
Laguna del Taray (Toledo) 15-20 - 6
Los Albardiales (Toledo) 2 - 0
Laguna de Vadancho (Toledo) 3-6 - 0
Lagunas de Villafranca (Toledo) 11 - 0
Laguna de El Masegar (Toledo) 15-19 - 0
Laguna de Pastrana (Toledo) 2-3 - 0
Prados de la Guardia (Toledo) 1 - 1
Manantial de Zampullones (Ciudad Real) 1 - 0
P. N. Las Tablas de Daimiel (Ciudad Real) 90-120 - 15
Embalse de Utxesa (Lleida) 15-25 - 22
Embalse de Camelis (Lleida) 1 - 0
P. N. Delta del Ebro (Tarragona) 50-100 - 68-80
P. N. Marjal de Pego-Oliva (Alicante-Valencia) 5 - 1
Prat de Cabanes-Torreblanca (Castellón) 0 - 1
P. N. s’Albufera (Mallorca) 12-13 - 21
Laguna de Las Cañas (Navarra) 1 - 6-8
Balsa del Juncal (Navarra) 10 - 0
Marisma de Alday (Cantabria) 3 - 0
Lagoa da Frouxeira (A Coruña) 3 - 5-6
Humidal de Sisalde-Barrañán (A Coruña) 3 - 1
Complexo litoral de Baldaio (A Coruña) 2 - 0
Lagoa de Vixán (A Coruña) 2 - 1
Esteiro do Ulla (A Coruña) 14-16 - 3
Esteiro do Miño (Pontevedra) 10 - 1
PORTUGAL (2015)
Ribeira de São Simão 1
Barrinha de Esmoriz Completo 7
Salreu (Parcial, 66%) 75-77
Gafanha Areão 1
Estuário do Tejo- Ponta da Erva 2
Estuário do Tejo -38 Moios 1
En 2005 se realizó el I Censo Nacional en España. Con resultado: E. s. witherbyi 250-360 parejas, la mayoría en Castilla-La Mancha; E. s. lusitanica 62-68 parejas, mayoría en Galicia.

Diez años después, en 2015, tras constatar un empeoramiento, se realizó con voluntarios el II Censo Nacional y I Censo Ibérico, coordinado por la Universidad de Valencia y la Universidad Politecnica de Valencia. El censo fue completo en España, pero no en Portugal, se visitaron 180 humedales (24 en Portugal, de los 31 potenciales), incluyendo todas las localidades censadas en 2005. En España, la población total es de 155-170 parejas (319-431 en 2005), con un declive del 50-60% en 10 años, por lo que la especie debería ser considerada en España y Portugal como En Peligro Crítico B2b (i,ii,iii,iv,v) según los criterios de la UICN.

El censo de escribano palustre iberoccidental fue de 96-101 parejas en 11 humedales, 5 en España (13 en 2015) y 6 en Portugal. En Portugal sólo se muestrearon el 77% de las localidades potenciales. La mayor parte se concentra en una localidad, Salreu, ría de Aveiro, Portugal (75-77 parejas). En España sólo hubo ejemplares en Galicia, con un censo de 11-12 parejas y un declive del 83% respecto a 2005 (un censo posterior, en 2018, localizó 20 parejas en Galicia). Para Portugal no existe un censo previo para comparar, aunque se detectó un descenso en el número de humedales con presencia como reproductor.
Para el escribano palustre iberoriental, el censo fue de 144-158 parejas reproductoras en 10 localidades (21 en 2005). El humedal con mayor población fue el Delta del Ebro (63-65%), seguido de Castilla-La Mancha (17-16%) y el P. N. de S’Albufera de Mallorca, única localidad de las islas Baleares (15-13%). Respecto a 2005 se ha detectado un declive del 43-56%, siendo más severo en las Tablas de Daimiel donde en 2005 se estimaron 90-120 parejas frente a las 15 detectadas en 2015. El escribano palustre iberoriental se reproduce en 2015 en sólo 10 humedales (21 en 2005) de Castilla-La Mancha, Cataluña, Comunidad Valenciana, Baleares y Navarra. En el litoral mediterráneo es estable en el Delta del Ebro, se reduce en la Comunidad Valenciana y aumenta en S’Albufera de Mallorca.

Portugal. Población reproductora en 2015 y método de censo. Universitat de València-Universitat
Politécnica de València-SEO/BirdLife. Madrid. https://doi.org/10.31170/0059
LOCALIDADES POSITIVAS EN ESPAÑA, 2005 y 2015
Laguna de los Ojos de Villaverde (Albacete) 7 - 0
Laguna de Manjavacas (Cuenca) 2 - 0
Embalse de Buendía-Río Mayor (Cuenca) 3 - 0
Complejo Lagunar de Arcas (Cuenca) 2 - 0
Embalse de Almoguera (Guadalajara) 7 - 3
Laguna del Taray (Toledo) 15-20 - 6
Los Albardiales (Toledo) 2 - 0
Laguna de Vadancho (Toledo) 3-6 - 0
Lagunas de Villafranca (Toledo) 11 - 0
Laguna de El Masegar (Toledo) 15-19 - 0
Laguna de Pastrana (Toledo) 2-3 - 0
Prados de la Guardia (Toledo) 1 - 1
Manantial de Zampullones (Ciudad Real) 1 - 0
P. N. Las Tablas de Daimiel (Ciudad Real) 90-120 - 15
Embalse de Utxesa (Lleida) 15-25 - 22
Embalse de Camelis (Lleida) 1 - 0
P. N. Delta del Ebro (Tarragona) 50-100 - 68-80
P. N. Marjal de Pego-Oliva (Alicante-Valencia) 5 - 1
Prat de Cabanes-Torreblanca (Castellón) 0 - 1
P. N. s’Albufera (Mallorca) 12-13 - 21
Laguna de Las Cañas (Navarra) 1 - 6-8
Balsa del Juncal (Navarra) 10 - 0
Marisma de Alday (Cantabria) 3 - 0
Lagoa da Frouxeira (A Coruña) 3 - 5-6
Humidal de Sisalde-Barrañán (A Coruña) 3 - 1
Complexo litoral de Baldaio (A Coruña) 2 - 0
Lagoa de Vixán (A Coruña) 2 - 1
Esteiro do Ulla (A Coruña) 14-16 - 3
Esteiro do Miño (Pontevedra) 10 - 1
PORTUGAL (2015)
Ribeira de São Simão 1
Barrinha de Esmoriz Completo 7
Salreu (Parcial, 66%) 75-77
Gafanha Areão 1
Estuário do Tejo- Ponta da Erva 2
Estuário do Tejo -38 Moios 1
domingo, 22 de abril de 2018



sábado, 21 de abril de 2018
BUSARDO RATONERO. POLIMORFISMO Y VARIACIÓN DE PLUMAJE
 |
| Busardo ratonero (Buteo buteo). Los siete tipos de plumaje utilizados en el Proyecto "Buteo Morph" |
El polimorfismo de color del plumaje se ha descrito en alrededor del 3,5% de las especies de aves, aunque las rapaces muestran una frecuencia mucho más alta, en especial el género Buteo con 15 especies polimórficas de 25. Estos polimorfismos son hereditarios y muy interesantes desde una perspectiva evolutiva. En la realidad se manifiestan como morfos discretos (águila calzada, halcón de Eleonora) o bien como una variación gradual continua (busardo ratonero, abejero europeo).
El poliformismo del busardo ratonero (Buteo buteo) ha sido bien estudiado en Europa, en especial en Holanda, donde un grupo de investigación recoge desde al menos 1996 más de 5.000 registros siguiendo un baremo con siete tipos de plumaje. El seguimiento de muchos individuos durante años indica que el color de plumaje se mantiene toda la vida del ave, en todo caso puede oscurecer un poco en el paso de edad juvenil a adulta. Los estudios clásicos solían definir tres morfos para Buteo buteo (oscuro, intermedio y claro), pero ahora se tienden a considerar siete categorías, desde muy oscuro a muy claro de modo gradual, en función de la proporción de color blanco en el plumaje. La asignación a un morfo determinado tiene cierta subjetividad y depende en parte de la luz y postura del ave.

Más información
http://aves.orn.mpg.de/~buteo/en
Kappers EF, de Vries C, Alberda A, Forstmeier W, Both C, Kempenaers B. 2018. Inheritance patterns of plumage coloration in common buzzards Buteo buteo do not support a one-locus two-allele model. Biol. Lett. 14: 20180007. http://dx.doi.org/10.1098/rsbl.2018.0007
Kappers, Elena Frederika; Bart Kempenaers y Christiaan Both. 2015. Discover colour variation in Common buzzards. Congreso EOU, Badajoz, agosto 2015.
Kappers E.F., Chakarov N., Krüger O., Mueller A.K., Valcu M., Kempenaers B. y Both C. 2017. Classification and temporal stability of plumage variation in Common Buzzards. Ardea 105: 125–136. doi: 10.5253/arde.v105i2.a1
 |
| Busardo ratonero. Polimorfismo. Proporción de los siete morfos establecidos en Holanda. Se aprecia la presencia de los siete morfos (en España no suele haber los dos morfos más claros), dominando las formas 2 y 3, es decir, oscura e intermedia-oscura) |
jueves, 19 de abril de 2018
FAMILIA LOCUSTELLIDAE. NUEVA TAXONOMÍA (2018)
La familia Locustellidae es una de las múltiples escisiones de la antigua gran familia Sylvidae. En la actualidad consta de 62
especies y 11 géneros. Alström et al (2018) analizan 59 especies y
modifican en gran medida la taxonomía previa. Los géneros Bradypterus,
Locustella, Megalurus, Megalurulus y Schoenicola no son monofiléticos. Tres de
los cinco géneros monotípicos (Amphilais, Buettikoferella y Malia) están dentro
de otros géneros, el género Chaetornis forma un clado con un Schoenicola. Se
propone la siguiente clasificación:

Alström, P., Cibois, A., Irestedt, M., Zuccon, D., Gelang, M., Fjeldså, J., Andersen, M.J., Moyle, R.G., Pasquet, E., Olsson, U. 2018. Comprehensive molecular phylogeny of the grassbirds and allies (Locustellidae) reveals extensive non-monophyly of traditional genera, and a proposal for a new classification. Molecular Phylogenetics and Evolution.
doi: https://doi.org/10.1016/j.ympev.2018.03.029

Alström, P., Cibois, A., Irestedt, M., Zuccon, D., Gelang, M., Fjeldså, J., Andersen, M.J., Moyle, R.G., Pasquet, E., Olsson, U. 2018. Comprehensive molecular phylogeny of the grassbirds and allies (Locustellidae) reveals extensive non-monophyly of traditional genera, and a proposal for a new classification. Molecular Phylogenetics and Evolution.
doi: https://doi.org/10.1016/j.ympev.2018.03.029
ÁGUILA PESCADORA. ALEMANIA. CICLO ANUAL
Águila pescadora (Pandion
haliaetus) en Alemania. Durante 1995-2011 se marcaron 28 adultos en el noreste
de Alemania con emisores satélite (duración hasta ocho años). Tres machos
invernaron en Iberia (una de cada ocho, el 12.5%) y 25 en África occidental. Las rutas migratorias en
Europa, especialmente en otoño, son más rectas y directas que en África
(posiblemente por el efecto del viento). Los machos en otoño no comienzan la
migración hasta terminar la cría cuando los pollos han volado. Las hembras
suelen migrar más temprano, antes de que los juveniles se independicen. Cuando
la reproducción fracasa, las hembras abandonan los nidos a finales de junio. El
inicio de la migración primaveral es similar en ambos sexos, aunque los machos
muestran poca variación en el momento de retorno y las hembras son más variables
(se supone que los machos compiten por los territorios y cuanto más temprano es
el regreso, más oportunidades tienen de conseguir un mejor territorio).
Bernd U. Meyburg, Dietrich Roepke, Christiane Meyburg, Andreas Bass, Rien E. van Wijk. 2018. The annual cycle of German adult Ospreys (Pandion haliaetus) – studies in the breeding and wintering areas as well as during migration since 1995 by means of satellite telemetry.
https://www.researchgate.net/project/Raptor-satellite-tracking-projects
Bernd U. Meyburg, Dietrich Roepke, Christiane Meyburg, Andreas Bass, Rien E. van Wijk. 2018. The annual cycle of German adult Ospreys (Pandion haliaetus) – studies in the breeding and wintering areas as well as during migration since 1995 by means of satellite telemetry.
https://www.researchgate.net/project/Raptor-satellite-tracking-projects
CULEBRA DE COLLAR (Género Natrix). NUEVA TAXONOMÍA (2018)
El género Natrix (Reptiles, Ofidios) constaba tradicionalmente de tres especies: culebra viperina Natrix maura (Mediterráneo occidental), Natrix tessellata (Mediterráneo oriental) y culebra de collar Natrix natrix (Paleártico). Recientes estudios de esta última han llevado a dividirla en tres especies distintas: Natrix astreptophora (Iberia y Magreb), Natrix helvetica (Europa centro-occidental e Italia) y Natrix natrix (Europa oriental y Asia occidental). También se ha descrito una cuarta especie: Natrix megalocephala (formas de cabeza grande en el Cáucaso), que Kindler et al (2018) descartan, por considerarla dentro de Natrix natrix.
En resumen, a fecha 2018 se aceptan las siguientes especies:
- Natrix maura - culebra viperina
- Natrix tessellata - culebra de dados
- Natrix astreptophora - culebra de collar ibérica
- Natrix helvetica - culebra de collar occidental
- Natrix natrix - culebra de collar oriental (incluye megalocephala)

Kindler, Carolin; Eva Graciá y Uwe Fritz. 2018. Extra-Mediterranean glacial refuges in barred and common grass snakes (Natrix helvetica, N. natrix). Scientific Reports, 8(1).
DOI:10.1038/s41598-018-20218-2
Kindler, Carolin; Chèvre, Maxime; Ursenbacher, Sylvain; Böhme, Wolfgang; Hille, Axel; Jablonski, Daniel; Vamberger, Melita y Fritz, Uwe. 2017. Hybridization patterns in two contact zones of grass snakes reveal a new Central European snake species. Scientific Reports, 7 (7378). doi:10.1038/s41598-017-07847-9
En resumen, a fecha 2018 se aceptan las siguientes especies:
- Natrix maura - culebra viperina
- Natrix tessellata - culebra de dados
- Natrix astreptophora - culebra de collar ibérica
- Natrix helvetica - culebra de collar occidental
- Natrix natrix - culebra de collar oriental (incluye megalocephala)
 |
| Natrix natrix. Subespecies antes de 2017. Con posterioridad se divide en tres especies: - Natrix astreptophora. Naranja. Península ibérica y noroeste de África - Natrix helvetica. Azules, violeta y verde claro. Centro-oeste Europa, Italia e islas mediterráneas - Natrix natrix. Amarillo, rojo y verde oscuro. Resto de Europa y Asia occidental |

Kindler, Carolin; Eva Graciá y Uwe Fritz. 2018. Extra-Mediterranean glacial refuges in barred and common grass snakes (Natrix helvetica, N. natrix). Scientific Reports, 8(1).
DOI:10.1038/s41598-018-20218-2
Kindler, Carolin; Chèvre, Maxime; Ursenbacher, Sylvain; Böhme, Wolfgang; Hille, Axel; Jablonski, Daniel; Vamberger, Melita y Fritz, Uwe. 2017. Hybridization patterns in two contact zones of grass snakes reveal a new Central European snake species. Scientific Reports, 7 (7378). doi:10.1038/s41598-017-07847-9
miércoles, 18 de abril de 2018
CULEBRILLAS CIEGAS IBÉRICAS. CAMBIOS EN LA NOMENCLATURA (2018)
Las culebrillas ciegas (género Blanus) de la Península Ibérica, tradicionalmente encuadradas en la especie Blanus cinereus, fue dividida en dos especies por Albert y Fernández (2009), que describen Blanus mariae sp. nov, distribuida por el suroeste peninsular y separada hace unos 5 millones de años. El tema ya fue tratado en el presente blog [enlace]. Sin embargo, Ceríaco y Bauer (2018) consideran que la descripción de Blanus mariae no es válida por un conjunto de errores, el principal que la localidad original para describir Blanus cinereus (Lisboa 1797) está en realidad dentro del área de distribución de Blanus mariae. Teniendo en cuenta morfología, genética e historia los autores revisan en un largo artículo de 33 páginas la nomenclatura de las culebrillas ciegas ibéricas proponiendo el mantenimiento de las dos especies citadas aunque con nuevas denominaciones: Blanus cinereus (se designa y describe un neotipo) y Blanus vandellii (se describe como especie nueva). Lo paradójico es que el nombre B. cinereus se aplica ahora a las poblaciones antes denominadas B. mariae distribuidas por el suroeste ibérico, mientras B. vandellii se aplica a la especie que ocupa el resto del territorio ibérico siempre denominada B. cinereus.
En Extremadura están presentes ambas especies, que parecen solapar en una parte importante del territorio aún por determinar. Así, el mapa adjunto indica la presencia de Blanus vandellii en dos puntos de la provincia de Cáceres (centro sur y Garciaz, en el sureste) y otros dos de la provincia de Badajoz (aparentemente entorno de Badajoz capital y de Jerez de los Caballeros). Por su parte Blanus cinereus está restringida al extremo sur de Badajoz (Oliva de la Frontera y Pallares), como ya se indicaba para "B. mariae", pero además aparece un punto de presencia en el centro de la región, más o menos en la sierra de Montánchez. No es posible precisar más, pues en la lista de localidades muestreadas no parecen figurar varias localidades extremeñas.
Luis M. P. Ceríaco y Aaron M. Bauer (2018) An integrative approach to the nomenclature and taxonomic status of the genus Blanus Wagler, 1830 (Squamata: Blanidae) from the Iberian Peninsula, Journal of Natural History, 52:13-16, 849-880.
DOI: 10.1080/00222933.2017.1422283
 |
Culebrillas ciegas (género Blanus) en la península Ibérica Puntos azules: Blanus cinereus (antes Blanus mariae) Puntos rojos: Blanus vandellii (antes Blanus cinereus) |
En Extremadura están presentes ambas especies, que parecen solapar en una parte importante del territorio aún por determinar. Así, el mapa adjunto indica la presencia de Blanus vandellii en dos puntos de la provincia de Cáceres (centro sur y Garciaz, en el sureste) y otros dos de la provincia de Badajoz (aparentemente entorno de Badajoz capital y de Jerez de los Caballeros). Por su parte Blanus cinereus está restringida al extremo sur de Badajoz (Oliva de la Frontera y Pallares), como ya se indicaba para "B. mariae", pero además aparece un punto de presencia en el centro de la región, más o menos en la sierra de Montánchez. No es posible precisar más, pues en la lista de localidades muestreadas no parecen figurar varias localidades extremeñas.
Luis M. P. Ceríaco y Aaron M. Bauer (2018) An integrative approach to the nomenclature and taxonomic status of the genus Blanus Wagler, 1830 (Squamata: Blanidae) from the Iberian Peninsula, Journal of Natural History, 52:13-16, 849-880.
DOI: 10.1080/00222933.2017.1422283
martes, 17 de abril de 2018
PERRO MAPACHE. EXPANSIÓN EN FRANCIA

El perro mapache (Nyctereutes procyonoides) es un cánido originario de Asia oriental (sobre todo China y Japón). Es una especie basal relacionada con los zorros del género Vulpes. Aunque en Asia está en declive, en Europa está en expansión y se considera invasora. Debido a su empleo en la industria peletera, la Unión Soviética soltó ejemplares por todo su territorio. Entre 1928 y 1958 se liberaron 10.000 ejemplares en 76 distritos de la URSS. En muchas regiones fracasaron, pero fueron exitosas en los estados bálticos, Ucrania y Rusia europea. En 1960 se cazaron en Letonia 4.210 ejemplares. Actualmente es abundante en Estonia, Finlandia, Letonia y Lituania, y se ha citado en Serbia, Francia, Rumania, Italia, Suiza, Alemania, Polonia, Noruega, Dinamarca y Suecia. Las poblaciones proceden de varios casos de introducción deliberada, así como de escapes de granjas.
Los siguientes mapas muestran la situación en Europa (año 2008) y en Francia, donde hay presuntas observaciones (sin pruebas) en los Pirineos, muy cerca de España.


CHACAL DORADO. EXPANSIÓN EN EUROPA

El chacal dorado (Canis aureus) se distribuye por el sur de Asia, con una distribución histórica marginal en Europa, donde ocupaba pequeñas zonas en el sureste (Balcanes) hasta mediados del siglo XX. Después de la II Guerra Mundial, quizás por la eliminación del lobo en grandes áreas, comenzó una expansión al oeste que aún continúa. La expansión parece que ha ocurrido en al menos tres oleadas: años 1950, años 1980 y principio del siglo XXI. Hasta 2015 se consideraban dentro del chacal dorado cánidos de la mitad norte de África. Varios estudios han demostrado que están más próximos al lobo común (Canis lupus) y ahora esta especie se denomina lobo dorado africano (Canis anthus), denominación dada en 1820 por Cuvier. Por tanto, 200 años después vuelve a su consideración original de especie diferenciada.
Desde finales de 2017 se han producido las primeras observaciones en Francia: al menos tres fotografías tomadas en Alta Saboya, límite con Suiza (donde se citó por primera vez en 2011). Como otros cánidos, por ejempo el zorro, el chacal es oportunista y puede vivir en cualquier hábitat que le ofrezca suficiente comida, incluyendo vertederos o zonas muy humanizadas. Estas citas en Francia se suman en la última década a la presencia de chacales dorados aislados en Países Bajos, Suiza, Dinamarca, Alemania, Letonia (2013), Lituania (2015) y Polonia (2015). En Austria (primera reproducción en 2007), noreste de Italia y Eslovenia su presencia parece ser estable. En Bulgaria, Rumania, Serbia y Croacia la población actual es muy numerosa (miles de ejemplares), en Grecia podría estar en declive y en Hungría el aumento es exponencial. Las limitaciones a su expansión son climáticas (evita zonas muy frías con nieves profundas en invierno) y ecológicas (presencia estable del lobo). La reacción a la colonización es variable entre países, así en Croacia es especie cinegética, en Alemania está protegido y en Lituania la quieren declarar especies invasora.
AMPLIACIÓN. La IUCN estima 117.000 ejemplares en Europa (¿2018?) y 17.000 lobos.
FUENTES
https://www.elperiodico.com/es/medio-ambiente/20180415/chacal-dorado-expansion-europa-llega-francia-6757820
https://www.ferus.fr/wp-content/uploads/2016/05/chacal-dore-europe-nathan-ranc.pdf
www.goldenjackal.eu
https://wilderness-society.org/golden-jackal-takes-on-europe/


 |
| Chacal dorado (Canis aureus). Distribución mundial y subespecies (Wikipedia). |
jueves, 12 de abril de 2018
MOSQUITEROS (familia Phylloscopidae). FILOGENIA
Nuevo estudio sobre la taxonomía de los mosquiteros (Aves: Phylloscopidae). Familia cuyo número de especies reconocidas ha aumentado un 50% en las últimas tres décadas (de 52 en 1980 a 76 especies en 2018), básicamente por estudios de vocalizaciones y ADN. Alström et al (2018) estudian todas las especies de mosquiteros del mundo y proponen una nueva filogenia. Datan el ancestro común más reciente en 11,7 millones de años (9,8-13,7 Ma), con línajes divergentes entre 0'5 Ma (0,3-0,8) y 6,1 Ma (4,8-7,5). Las especies del tradicional género Seicercus quedan todas encajadas dentro del género Phylloscopus, que pasaría a ser el único género de la familia Phylloscopidae (criterio seguido en HBW Alive). El área de distribución es el Viejo Mundo, en especial el centro de Asia (16 especies en los Himalayas orientales y 20 especies en los montes Qinling, norte-centro de China).
Alström, P., Rheindt, F.E., Zhang, R., Zhao, M., Wang, J., Zhu, X., Yin Gwee, C., Hao, Y., Ohlson, J., Jia, C., Prawiradilaga, D.M., Ericson, P.G.P., Lei, F., Olsson, U. 2018. Complete species-level phylogeny of the leaf warbler (Aves: Phylloscopidae) radiation, Molecular Phylogenetics and Evolution.
doi: https://doi.org/10.1016/j.ympev.2018.03.031

Alström, P., Rheindt, F.E., Zhang, R., Zhao, M., Wang, J., Zhu, X., Yin Gwee, C., Hao, Y., Ohlson, J., Jia, C., Prawiradilaga, D.M., Ericson, P.G.P., Lei, F., Olsson, U. 2018. Complete species-level phylogeny of the leaf warbler (Aves: Phylloscopidae) radiation, Molecular Phylogenetics and Evolution.
doi: https://doi.org/10.1016/j.ympev.2018.03.031

martes, 10 de abril de 2018



LOBO. FRANCIA. EXPANSIÓN 2002-2017
Lobo en Francia (población):
- años 1930: extinción del lobo en Francia
- 1992: se instalan dos ejemplares procedentes de Italia que fundan la primera manada moderna. Con nuevos ejemplares de Italia se establecen en el P. N. Mercantour y se extienden al norte (Alpes) y mucho más tarde al sur (cruzan el Ródano y llegan al Macizo Central y los Pirineos)
- 2008: 194 ejemplares en invierno
- 2012: 250 ejemplares en invierno
- 2013: 119-229 ejemplares en marzo
- 2017: 265-402 ejemplares en marzo
_______________________________________________________________
El Gobierno francés ha presentado el plan de gestión del lobo. Entre otras cosas prevé matar un máximo de 40 ejemplares en un año para limitar los daños al ganado. La población en marzo de 2017 se estima en 265-402 lobos (aumento respecto a 119-229 en marzo de 2013) y unas 50 manadas. Las indemnizaciones por valor de 3,2 millones de euros en 2016 (+60% respecto a 2013). Matar un lobo en Francia ilegalmente es un delito penado con hasta dos años de cárcel y 150.000 euros de multa.
http://www.eldiario.es/cultura/tecnologia/Francia-autoriza-matar-limitar-ataques_0_741926289.html
_____________________________________________________________
En el periodo 2008-2012 el área de ocupación ha aumentado un 25% y la población estimada de 194 lobos en el invierno 2008-2009 a 250 ejemplares en el invierno 2012 (según método de captura-marcado-recaptura). Se ha pasado de 25 zonas de ocupación permanente en 2008 (16 manadas) a 29 en 2012 (19 manadas).
- años 1930: extinción del lobo en Francia
- 1992: se instalan dos ejemplares procedentes de Italia que fundan la primera manada moderna. Con nuevos ejemplares de Italia se establecen en el P. N. Mercantour y se extienden al norte (Alpes) y mucho más tarde al sur (cruzan el Ródano y llegan al Macizo Central y los Pirineos)
- 2008: 194 ejemplares en invierno
- 2012: 250 ejemplares en invierno
- 2013: 119-229 ejemplares en marzo
- 2017: 265-402 ejemplares en marzo
_______________________________________________________________
El Gobierno francés ha presentado el plan de gestión del lobo. Entre otras cosas prevé matar un máximo de 40 ejemplares en un año para limitar los daños al ganado. La población en marzo de 2017 se estima en 265-402 lobos (aumento respecto a 119-229 en marzo de 2013) y unas 50 manadas. Las indemnizaciones por valor de 3,2 millones de euros en 2016 (+60% respecto a 2013). Matar un lobo en Francia ilegalmente es un delito penado con hasta dos años de cárcel y 150.000 euros de multa.
http://www.eldiario.es/cultura/tecnologia/Francia-autoriza-matar-limitar-ataques_0_741926289.html
_____________________________________________________________
En el periodo 2008-2012 el área de ocupación ha aumentado un 25% y la población estimada de 194 lobos en el invierno 2008-2009 a 250 ejemplares en el invierno 2012 (según método de captura-marcado-recaptura). Se ha pasado de 25 zonas de ocupación permanente en 2008 (16 manadas) a 29 en 2012 (19 manadas).
Fuente: Ministerio de Medio
Ambiente de Francia (2013). Projet de Plan d’action national loup 2013-2017.
 |
| Lobo. Francia. Expansión. Mapas 1992, 1997, 2002, 2007 y 2011 |
 |
| Lobo. Francia. 2012. Áreas de ocupación permanente n=29 (dos Pirineos, 27 Alpes y pre-Alpes) |
CORZO. EXPANSIÓN EN ESPAÑA (1969-2007)
 |
| Corzo. Distribución en España en 1969. |
 |
| Corzo. Expansión en España entre 1969 y 2002 (algunas poblaciones tienen su origen en repoblaciones, como el caso de Las Batuecas, Salamanca). |
 |
| Corzo. Distribución según el Atlas de España, año 2001 |
 |
| Corzo. Distribución en España según el Atlas de 2007. Este atlas no es un nuevo estudio, sino una ampliación del atlas de 2001, que en el caso del corzo sólo se traduce en una mayor área de ocupación en algunas provincias, más evidente en País Vasco, Cáceres y sur de Castilla-La Mancha. En el caso de Cáceres, el área de ocupación se considera exagerada y por encima de la real (al parecer los datos son estimaciones de ocupación a partir del hábitat). |
lunes, 9 de abril de 2018
ALCAUDÓN DORSIRROJO. DECLIVE DEL 95% EN EL PAÍS VASCO (1974-2016)
José Luis Tellería (2018), tras analizar dos poblaciones de alcaudón dorsirrojo en Vizcaya a lo largo de cuatro décadas, determina un declive del 95% entre 1974 y 2016. Las localidades estudiadas son Ea (campiña costera) y Landa (pastos y brezales de interior). La primera baja de 24 y 21 parejas en 1974 y 1975 a solo una pareja en 2014 y ninguna en 2016. La segunda cayó de 22 en 1974 y 15 parejas en 1975 a sólo dos en 2014 y una en 2016. Entre ambas, pasan de 41 a 1 parejas (-95% en 42 años).
Por tanto, el declive es anterior a la puesta en marcha del programa SACRE en 1998, que detecta caídas del 58% (1998-2015), del 75% (1998-2016) y del 69% (1998-2017) en el País Vasco. En el conjunto de España también muestra una tendencia negativa del 50% entre 1998 y 2017. Cifras que contrastan con la evolución a escala europea, donde los datos de EBCC indican estabilidad.


No es posible determinar las posibles causas. Las plantaciones forestales ocasionan la pérdida de 21 territorios, al ocupar el 20-40% del hábitat inicial. Ha habido proliferación urbana en Ea pero no en Landa. Y las prácticas agrarias relacionadas con la PAC tienen escaso efecto (se utilizan pocos biocidas).
Referencia:
José Luis Tellería. 2018. Viejos conteos sugieren el colapso de dos poblaciones de alcaudón dorsirrojo Lanius collurio. Ardeola, 65(2). [Nota de prensa: https://www.seo.org/2018/04/05/alarmante-declive-del-alcaudon-dorsirrojo-en-el-pais-vasco/]
Por tanto, el declive es anterior a la puesta en marcha del programa SACRE en 1998, que detecta caídas del 58% (1998-2015), del 75% (1998-2016) y del 69% (1998-2017) en el País Vasco. En el conjunto de España también muestra una tendencia negativa del 50% entre 1998 y 2017. Cifras que contrastan con la evolución a escala europea, donde los datos de EBCC indican estabilidad.


No es posible determinar las posibles causas. Las plantaciones forestales ocasionan la pérdida de 21 territorios, al ocupar el 20-40% del hábitat inicial. Ha habido proliferación urbana en Ea pero no en Landa. Y las prácticas agrarias relacionadas con la PAC tienen escaso efecto (se utilizan pocos biocidas).
Referencia:
José Luis Tellería. 2018. Viejos conteos sugieren el colapso de dos poblaciones de alcaudón dorsirrojo Lanius collurio. Ardeola, 65(2). [Nota de prensa: https://www.seo.org/2018/04/05/alarmante-declive-del-alcaudon-dorsirrojo-en-el-pais-vasco/]
jueves, 5 de abril de 2018
PALOMA TORCAZ. IBERIA. MIGRACIÓN E INVERNADA
 |
Paloma torcaz. Migración e invernada en la península Ibérica. Con flechas se marcan las principales vías de paso migratorias, en azul las entradas por el Pirineo y en rosa los movimientos dentro de la península (suelen ser collados en montaña). El área azul muestra la vía principal de migración en una amplia franja al norte del Sistema Central. Los círculos naranjas son las principales áreas de invernada. Fuente: www.torcaces.com |
ÁGUILA REAL. DISPERSIÓN DE UN EJEMPLAR MARCADO EN ORENSE

Desplazamiento de un ejemplar de águila real (Ana) marcada por GREFA y liberado en la provinica de Orense. Dispersa hacia la provincia de Cáceres, donde permanece en varias zonas antes de regresar a Galicia.

LAGARTIJA BATUECA. REVISIÓN

Distribución: sierra de Francia, las Batuecas y las Mestas, en el sur de Salamanca y, marginalmente, extremo norte de Cáceres (únicamente conocida en el pico Rongiero, Sierra de La Granjera, Las Hurdes). Ocupa el polígono formado por la Peña de Francia, Hastiala y Rongiero, más una pequeña zona en el Portillo-Pico Orconera. Carbonero et al (2016) realizaron un estudio exhaustivo en abril-octubre de 2007 y 2008 por encargo del Gobierno de Castilla y León. Muestrearon un área potencial de 63 cuadrículas UTM 1x1 km. Sólo detectado en 23 cuadrículas UTM 1x1 km (36%) y tres cuadrículas UTM 10x10 km, área total de 20-25 km2, área real de 12-15 km2, repartida de modo discontinuo en núcleos dispersos en pedrizas. Se diferencian dos zonas independientes. (1) Noroeste del Parque Natural de Las Batuecas-Sierra de Francia, en altitudes de 886-1.730 m, limitada por los picos Peña de Francia (1.723 m), Robledo (1.611 m), Hastiala (1.735 m), Alto del Copero (1.560 m), Mesa del Francés (1.640 m) y Rongiero (1.627 m); en esta zona se ha localizado en 22 cuadrículas de 1x1 km en pedrizas de cuarcita y granito. (2) Segunda zona en laderas meridionales de las sierras de La Alberca y el Puerto del Portillo, en una única cuadrícula de 1x1 km y altitudes de 840-1.400 m. La existencia de este pequeño núcleo hace posible la presencia de otros núcleos sin descubrir, incluso al oeste, en la sierra de Gata, donde ningún estudio ha podido aún confirmarlo.
 |
Cuadrículas de 1x1 km del Parque Natural de las Batuecas y Sierra de Francia. En gris la zona prospectada: en gris oscuro las cuadrículas con presencia de lagartija batueca y en gris claro las cuadrículas donde no se detectó la especie. |
Hábitat: canchales estables de grandes rocas con abundancia de musgos y líquenes en pendientes moderadamente inclinadas. Pisos meso, supra y oromediterráneo entre 840 y 1.730 m. En el mesomediterráneo en canchales entre bosques de encina (Quercus ilex) y alcornoques (Quercus suber); en el supramediterráneo entre bosques de robles melojos (Quercus pyrenaica) y en oromediterránea (cumbres de Peña de Francia y pico Hastiala) entre piorno serrano.
Población: se estiman 1.200-1.500 ejemplares. En Peña de Francia (1.600-1.723 m) se estiman 30 ejemplares (densidad 10 ej/ha). Las poblaciones de cumbres son pequeñas y no suponen el grueso de la población, que está en zonas más bajas del valle de las Batuecas. En el Portillo (840-1.400 m), densidades de 25-30 ej./ha con máximos de 58 ej./ha. Densidad media para toda su área de distribución 24 ej./ha (unas 10 veces inferiores a otras Iberolarceta del Sistema Central, como I. cyreni e I. monticola). Muy pocos datos de tendencia de la población, al menos con declive en la Peña de Francia.
Conservación: "En Peligro Crítico" según criterios UICN. Se solicita "En peligro de extinción" en el catálago español, donde actualmente figura en el LESRPE.
Extremadura: ¿presencia? Alberto Gil en 2014 (http://desdeeltorreon.blogspot.com.es/2014/05/la-lagartija-de-las-batuecas.html) afirma que "sí se ha encontrado en Las Hurdes (Cáceres), aunque en un área tan reducida que, si tenemos en cuenta las densidades obtenidas en otras zonas para la especie, no creo que sean más de 20-50 individuos". Por su parte, Óscar Arribas (2015) publica lo siguiente: "...está presente también en la cara norte de la Sierra de las Mestas (pico Rongiero, 1.627 m)... Muy probablemente habite también en la vertiente sur, perteneciente ya a la Comunidad Extremeña", por tanto no confirma claramente esta presencia.
BIBLIOGRAFÍA
Carbonero, J.; García-Díaz, P.; Ávila, C.; Arribas, O. y Lizana, M. 2016. Distribution, habitat characterization and conservation status of Iberolacerta martinezricai (Arribas, 1996), in the Sierra de Francia, Salamanca, Spain. Herpetozoa, 28:149-165.
Arribas, O. 2015. Lagartija batueca - Iberolacerta martinezricai (Arribas, 1996). En: Carrascal, L. M. & Salvador, A. (eds.). Enciclopedia Virtual de los Vertebrados Españoles. Museo Nacional de Ciencias Naturales. Madrid. http://www.vertebradosibericos.org/
ANEXO
 |
| Resultados por localidades obtenidos por Carbonero et al (2016) durante un estudio exhaustivo realizado en abril-octubre de 2007 y 2008 por encargo del Gobierno de Castilla y León |
CIGÜEÑA BLANCA. PORTUGAL. NIDOS EN TENDIDOS DE ALTA TENSIÓN

Francisco Moreira, Ricardo C. Martins, Inês Catry y Marcello D'Amico. 2018. Drivers of power line use by white storks: a case study of birds nesting on anthropogenic structures. Journal of Applied Ecology. https://doi.org/10.1111/1365-2664.13149
CARRICERÍN CEJUDO. ANILLAMIENTO EN FRANCIA. 2013-2016
 |
Carricerín cejudo. Francia. Número de ejemplares capturados para anillamiento (1969-2016) |
 |
| Carricerín cejudo. Francia. Fenología diaria de paso postnucpcial año 2016. |
 |
Carricerín cejudo. Francia. Fenología mensual de paso postnupcial de adultos y juveniles. |
Hemery D., Blaize C., Musseau R. y Jiguet F. 2017. Les captures de Phragmites aquatiques en France, 2013-2016. Plan national d'actions en faveur du Phragmite aquatique. Dreal Bretagne – Bretagne Vivante-SEPNB. 22p.
CARRACA EUROPEA. MIGRACIÓN DE LA POBLACIÓN IBÉRICA
Las carracas ibéricas utilizaron al menos dos rutas migratorias alternativas durante la migración postnupcial, una directa hacia el sur desde el noreste ibérico y un desvío por la costa atlántica africana desde las poblaciones
del sur, en lo que parecen dos estrategias diferentes para afrontar el cruce del desierto del Sáhara. La zona del Sahel, concretamente los alrededores del Lago Chad, es un área clave de descanso o escala, mientras que Angola, Botswana y Namibia constituyen el área principal de invernada.

Los individuos marcados mostraron una conectividad migratoria débil (poca relación entra áreas de cría y de invernada), pues todas las aves ibéricas confluyen en invierno en un área restringida, confiriendo una alta vulnerabilidad ante potenciales cambios ambientales. El desarrollo de modelos de distribución indican que las zonas de invernada de las carracas ibéricas suponen una pequeña porción al noroeste del área total estimada para la especie, ocupando áreas con un grado de protección mayor.

Rodríguez-Ruiz, J. 2017. Selección de hábitat y ecología del movimiento en un migrante transahariano: Contribución a la conservación de la Carraca europea. Ecosistemas 26:121-125.
DOI: 10.7818/ECOS.2017.26-1.19

Los individuos marcados mostraron una conectividad migratoria débil (poca relación entra áreas de cría y de invernada), pues todas las aves ibéricas confluyen en invierno en un área restringida, confiriendo una alta vulnerabilidad ante potenciales cambios ambientales. El desarrollo de modelos de distribución indican que las zonas de invernada de las carracas ibéricas suponen una pequeña porción al noroeste del área total estimada para la especie, ocupando áreas con un grado de protección mayor.

Rodríguez-Ruiz, J. 2017. Selección de hábitat y ecología del movimiento en un migrante transahariano: Contribución a la conservación de la Carraca europea. Ecosistemas 26:121-125.
DOI: 10.7818/ECOS.2017.26-1.19
LAGARTIJAS CENICIENTA, DE EDWARDS Y OCCIDENTAL IBÉRICA (Complejo Psammodromus hispanicus). DISTRIBUCIÓN
 |
Lagartija occidental ibérica (Psammodromus occidentalis). Oliva de Plasencia, Cáceres. (Hugo Sánchez) |
La lagartija cenicienta (tradicionalmente Psammodromus hispanicus en sentido amplio) fue diferenciada en el año 2010 (Fitze et al 2010) en tres especies distintas. Al ser especies crípticas muy similares entre sí, aunque diferenciables morfológicamente por expertos, las áreas de distribución de las tres no están bien definidas. El siguiente mapa muestra las áreas seguras para las tres especies, no siendo posible establecer la distribución en algunas zonas, sobre todo en la franja central norte-sur de la península Ibérica.

http://www.vertebradosibericos.org/reptiles/listareptiles.html
Fitze, P. S., González Jimena, V., San José García, L. M., San Mauro, D., Aragón, P., Suárez, T., Zardoya, R. 2010. Genetic, ecological, phenotypic and geographic diversity in the Psammodromus hispanicus species group. Pp. 364. En: Andreu, A. C., Beltrán, J. F., Tejedo, M. (Eds.) XI Congresso Luso-Espanhol de Herpetologia.

{kind=link}
lunes, 2 de abril de 2018
ESPÁTULA COMÚN. POBLACIÓN NIDIFICANTE EN PAÍSES BAJOS Y FRANCIA. DATOS ACTUALES
 |
| Espátula comun. Área de reproducción en Europa (Wikipedia). Faltan, al menos, las colonias del interior de la península Ibérica (Portugal y Extremadura) y sobra el Delta del Ebro |
La población de espátula común (Platalea leucorodia) en Europa occidental
se reparte entre los Países Bajos (cerca de 3.000 parejas en Holanda, pero
también cría en Alemania, Dinamarca, Reino Unido y Bélgica), Francia (562-618 pp en 2012) y España (>1.500
parejas con fuertes fluctuaciones). La población de Europa oriental ocupa Italia, Austria, Hungría, Chequia, Balcanes, Mar Negro, Grecia y sur de Ucrania.
En Francia, la población ha aumentado desde 12 parejas en 1992 a 618 en 2012. Anida en Somme, Loire-Atlantique, Charente-Maritime, Landes y Bouches-du-Rhone. Las principales colonias están en Camargue, Bouches-du-Rhone (143 parejas en 2011), y Lac Grand-Lieu, Loire-Atlantique (95 parejas en 2012). Invernan principalmente en la Península Ibérica y en África occidental, pero también en las costas del oeste y sur de Francia (940 aves, enero de 2012).
En los Países Bajos aumentó desde 148 parejas en 1968 a 2.534 en 2012 (38
colonias, el 60% en las islas de Frisia, especialmente Texel) y cerca de 3.000 en 2015. En 2015, la
colonia de Geul, la más grande del país, tenía 641 nidos (récord). La especie se
estableció en Texel después de 1900, en Terschelling en 1962, en Vlieland en
1983, en Schiermonnikoog en 1992, en Ameland en 1996, en Rottumerplaat en 1998 y
en Griend en 2010. En el delta del Rhin, se estableció a finales de los 1980, con un aumento muy
notable y unas 200 parejas en la actualidad. Las Islas Frisias suman unos 1.500 nidos después de 30 años de crecimiento exponencial.
 |
Espátula comun nidificante en Holanda. Mapa con las colonias y gráfica con el número de nidos entre 1960 y 2015 |
 |
| Espátula comun nidificante en Holanda. Gráfica de censos entre 2001 y 2013 |
En 2015, tres colonias del mar de Wadden sumaban 372 parejas, con un éxito
reproductivo entre 0,5 pollos/pareja en la Isla de Griend y 1,56 pollos/pareja
en Ameland, por debajo de los 2 pollos/pareja de la década de 1990. Esto se
achaca a la saturación poblacional y por tanto a la mayor competencia por el
alimento. Al comparar diez colonias se concluye que la tasa de supervivencia de
los pollos y el número de crías por nido eran más bajos en colonias estables que
en colonias en aumento. En 1991 y 2011 se encontró una clara correlación
negativa entre éxito reproductivo y tamaño de la colonia. Esta situación explica
la expansión en el delta del Rin y las islas Wadden en Alemania y Dinamarca (en
Schleswig-Holstein, Alemania, han aumentado un 25% sólo en una temporada, entre
2016 y 2017). Es probable que las colonias francesas también se estén
beneficiando de la expansión desde Holanda.
 |
| Espátula común. Holanda. Islas frisias que acogen las principales colonias y donde se han marcado la mayor parte de las aves que se ven en migración e invernada en la península Ibérica. |
Suscribirse a:
Entradas (Atom)